Two Frontal Hubs, Two Attention Pathways: How Do FEF and IFJ Tune the Brain with diverging oscillations?
Prompt to inhance my writing:
Mistakes found in the Veneiro Paper:
Figure S8: “frequency bands illustrated in Fugure 4 !!! not 5”
Figure 3: FEF, IFJa and IFJp not FEF!!!
ToDo-List
- theta reset quellen finden
- Quelle finden wo Theta auf dem PFC kommt
- Interesting point to add: there could be cross frequency coupling between beta and theta waves in IFJp
- [IFJ distinction between IFJa und IFJp
my text so far 15.06. 19:30
\begin{abstract}
While the classic “what” vs. “where” dichotomy in visual attention is well-supported anatomically, less is known about how these streams avoid interference in time. Building on the Communication Through Coherence (CTC) framework, this paper reviews evidence that the Frontal Eye Field (FEF) and Inferior Frontal Junction (IFJ) differentially control dorsal and ventral visual pathways via distinct oscillatory frequencies. I propose a “Clock-and-Data” hypothesis, where IFJp acts as a theta-rhythmic pacemaker to time-separate beta- and delta-band routing of spatial and object-based attention.
\end{abstract}
\section{Introduction}
Selective Attentional routes have long been differentiated into “what” and “where” streams. But still the question remains of how these streams interfere or stay out of each other’s way.
The classic dichotomy of the streams shows attention being top-down through low-frequencies while sensory-driven activity in higher frequencies \cite{buschman_top-down_2007} while Duncan and Desimone have framed attention as a biased competition but left timing unspecified \cite{duncan_neural_nodate}.
In Fries 2015, his Communication Through Coherence Hypothesis states that two neural groups can exchange information efficiently only when they rhythmically phase-align with each other. Coherence is like a window-opener of the upper region giving the opportunity for input. The sender that matches exactly this time window entrains the higher region and couples both of them \cite{fries_rhythms_2015}.
If they are miss-aligned, the same anatomical connection becomes silent. This shows the importance of phasic-coupling for cortical communication.
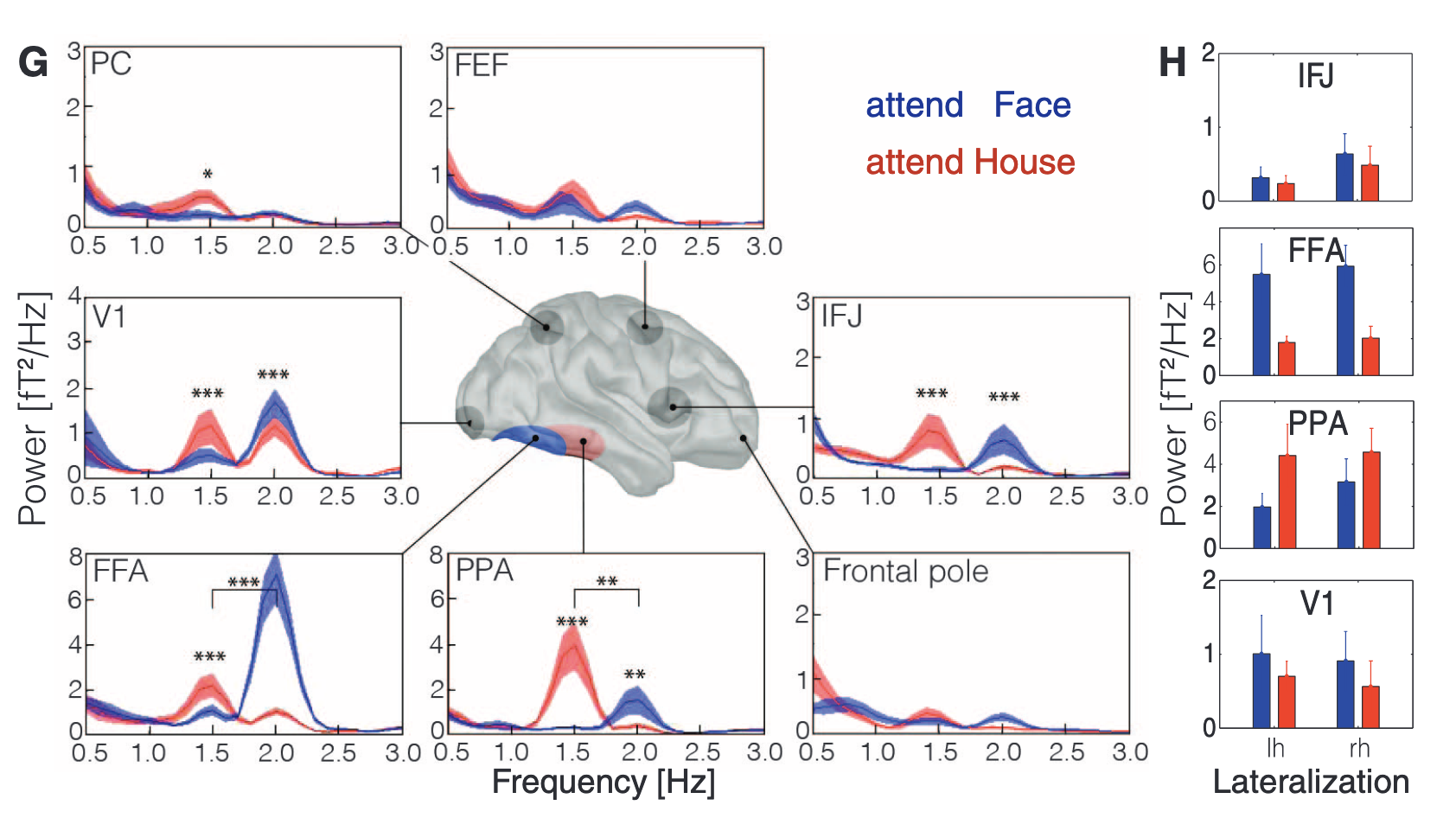
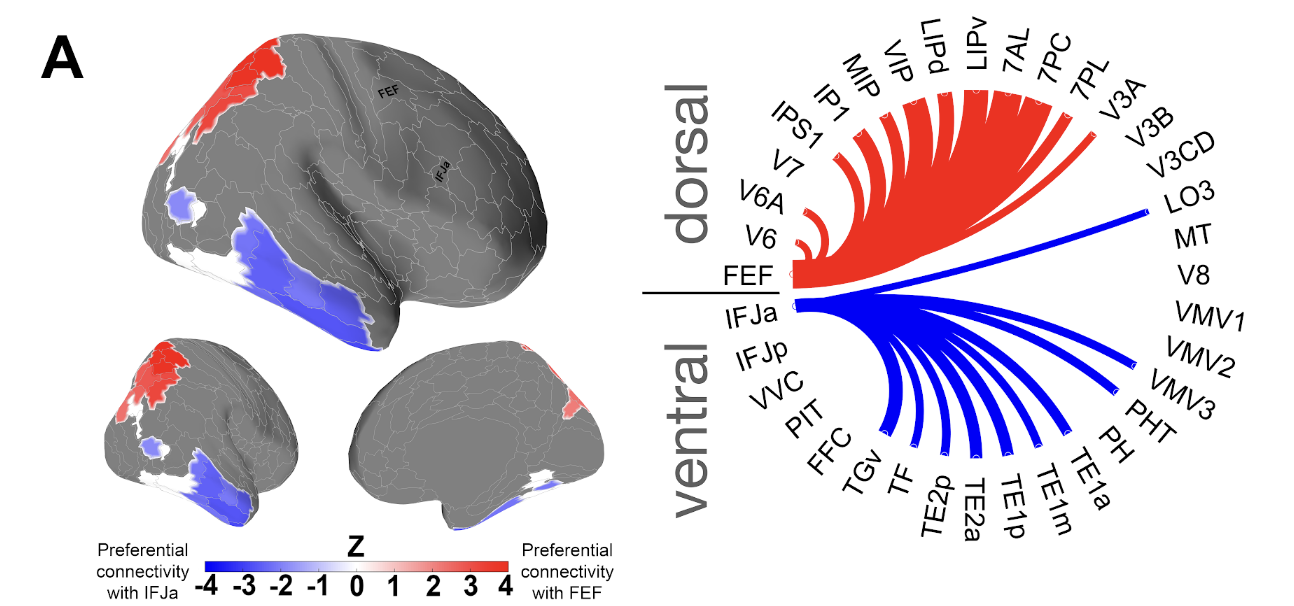
In the paper of Baldauf & Desimone 2014, they proof IFJ(Inferior Frontal Junction) being the synchronous attentional driver of the ventral stream \cite{baldauf_neural_2014}. While a paper from 2022 shows proof that connections from FEF(Frontal Eye Field) are mostly signifantly connected to dorsal regions while IFJ is connected to ventral regions \cite{soyuhos_functional_2022}.
These studies show which regions talk to each other, but do not mention when this communication is happening.
Looking at temporal encoding matters, as Fries stated, so the two pathways with FEF-dorsal and IFJ-ventral should not interfere with each other at the same frequency or timing,
Otherwise these two would collide on shared coritcal territory.
Spatial attention travels on a beta-band-block from FEF to dorsal \cite{soyuhos_functional_2022} while object-featured attention travels on a delta-band. Also IFJp sends infromation both to the IFPa and FEF, maintaining at theta oscillations. Also according to Fries, slower theta oscillations server as a pacemaker for lower regions, resetting when the gaze shifts.
This suggests, IFJp could serve as a pacemaker while then both FEF and IFJa receive the different oscillations for their resepctive streams and therefore do not interfere.
The IFJp fulfills ctriterias as a pacemaker. \href{https://www.overleaf.com/learn}{help library}, or head to our plans page to \href{https://www.overleaf.com/user/subscription/plans}{choose your plan}.
\section{Do FEF and IFJa show divergent intrinsic connectivity profiles?}
Looking at the band-width at which FEF and IFJa communicate to its corresponding regions we see the follwing.
\refstepcounter{figure}
\begin{center}
\noindent
\includegraphics[width=0.8\columnwidth]{localizationOfFEF.png}
\vspace{4pt}
\captionof{figure}{%
Localization of FEF and IFJ through an fMRI meta-analysis by \cite{bedini_accurate_2023}, showing that only the small region 55b is located between FEF and IFJ. Label as defined according to the MMP1 atlas.}
\label{fig:localization-of-FEF}
\end{center}
This is normal text: FEF always different frequency than IFJa?
\subsection{FEF mediates top-down attention}
FEF is known for its important role in attention in the dorsal-visual-stream. Through TMS the FEF is stimulated and therefore selectively phase-aligns in the beta-band with occipital sites, while no other frequency showed phase consistency \cite{veniero_top-down_2021}.
\textcolor{blue}{attentional top-down control signals by brief FEF stimulation do not only phase-reset oscillatory activity in the beta-band but also caused visual performance to cycle at the same frequency. from \cite{veniero_top-down_2021}}
\section{Clock-and-Data Hypothesis}
As Fries states in his paper, theta-oscillations serve as a setback for lower coritcal regions. As when attention shifts, it happens in a theta-rhythm.
\begin{center}
\centering
\includegraphics[width=0.8\linewidth]{theta-oscillation-reset.png}
\captionof{figure}{ \textcolor{red}{Überarbeiten!!}Theta-Rhythmic Phase Reset Turns Gamma-Frequency Differences into Latency Differences.
In the lower area, after a reset, the gamma rhythms representing different stimuli start at the same phase.
The gamma rhythm representing the attended stimulus (green) is faster than the gamma rhythm representing the unattended stimulus (orange).
This frequency difference translates into a latency difference: input from the attended representation reaches the higher area first,
transmits its representation, and triggers inhibition to shut out the competing unattended representation.}
\label{fig:theta-oscillation-reset.png}
\end{center}
I propose attention is based on time-devision and serves a intertwined oscillation pattern of theta serving as the clock and reset by the IFJp and the beta- and delta-oscillations serve as the data transfer from IFJa to ventral stream and FEF to dorsal stream.
Two frontal attentional hubs: Do FEF and IFJa show divergent intrinsic connectivity profiles?
- FEF always different frequency than IFJa?
Are oscillations frequency-specific to spatial vs. object attention?
- beta with FEF
- delta with IFJ
How is it transferrable to audition?
Chatty
Abstract
While the classic “what” vs. “where” dichotomy in visual attention is anatomically well-supported, less is known about how these streams avoid temporal interference. Building on the Communication Through Coherence (CTC) framework, this paper reviews evidence that the Frontal Eye Field (FEF) and Inferior Frontal Junction (IFJ) differentially control dorsal and ventral visual pathways via distinct oscillatory frequencies. I propose a “Clock-and-Data” hypothesis, in which a shared theta rhythm, originating from the prefrontal cortex (PFC), serves as a pacemaker that time-separates beta- and delta-band routing of spatial and object-based attention. This hypothesis not only integrates multiple findings from neuroimaging and electrophysiology, but also provides a testable framework for future studies investigating the rhythmic basis of attention.
1. Introduction
Selective attention has long been divided into “what” and “where” pathways. But how do these streams avoid interfering with each other in time? The classic model posits that top-down attention is conveyed via low-frequency oscillations, while bottom-up signals use higher frequencies (Buschman & Miller, 2007). Duncan and Desimone’s biased competition model adds another layer by emphasizing competitive selection, yet remains vague on timing mechanisms (Desimone & Duncan, 1995).
Fries’ (2015) CTC hypothesis offers a compelling solution: communication between neural populations is only effective when their rhythms are phase-aligned. A misaligned sender cannot effectively transmit, regardless of structural connectivity. This highlights temporal coherence as a prerequisite for functional coupling.
Evidence supports that FEF and IFJ engage distinct visual pathways at different frequencies. FEF couples with dorsal stream areas (e.g., SPL) via beta rhythms, supporting spatial attention (Veniero et al., 2021). Conversely, IFJ preferentially couples with ventral visual regions in the delta and gamma bands, facilitating object-based attention (Baldauf & Desimone, 2014).
But this anatomical and frequency-based divergence invites a deeper question: **how are these frequencies coordinated?**Here, the “Clock-and-Data” hypothesis emerges: a shared theta rhythm may periodically reset both pathways, allowing beta and delta bands to operate in separate windows. This rhythmic time-division could prevent cross-talk even within overlapping cortical regions.
2. Divergent Connectivity of FEF and IFJ
Figure 1 illustrates the anatomical separation of FEF and IFJ, with area 55b as the key boundary (Bedini et al., 2023). Resting-state fMRI reveals that FEF exhibits strong beta coupling with dorsal visual areas, whereas IFJ aligns more with ventral regions at lower frequencies.
2.1 FEF: Top-Down Spatial Attention
The FEF, located at the posterior dorsolateral prefrontal cortex (pDLPFC), directs spatial attention through beta-band coupling with dorsal stream regions. TMS studies confirm that FEF entrains occipital areas in the beta band, while other frequencies show no consistent effects (Veniero et al., 2021). Phase-resetting in the beta band enhances visual cortex excitability, particularly V5.
This points to a stable communication mode: spatial attention is routed from FEF to dorsal regions through beta oscillations. This finding is consistent with the notion that spatial attention must be fast, sustained, and spatially precise—characteristics that align well with beta-band dynamics. Additionally, beta-band coupling is often linked to the maintenance of cognitive states, suggesting that the FEF may act as a stabilizer of attentional focus once a spatial target is selected.
2.2 IFJ: Feature and Object-Based Attention
The IFJ shows a functional division: IFJa supports feature-based attention via delta and gamma rhythms, while IFJp interacts with both IFJa and FEF, particularly in the theta band (Veniero et al., 2021).
-
IFJa: Phase-couples with inferior/middle temporal cortex in delta (1–4 Hz) and gamma (30–100 Hz) bands.
-
IFJp: Exhibits stronger alpha/beta synchrony with FEF, but weaker links to ventral areas.
Multivariate pattern analyses (Meyyappan et al., 2021) show IFJa carries feature-selective information, while IFJp encodes mixed spatial and feature signals—suggesting a relay role.
Notably, IFJp may not serve as the pacemaker itself, due to lack of dominant theta activity. Instead, we posit that it relays a theta rhythm from a more anterior PFC region, possibly area 10. This relay mechanism maintains timing while allowing IFJp to gate downstream routing selectively.
This implies a hierarchy not only in anatomical terms but in temporal control. The PFC may operate as a clock, while IFJp functions as a switchboard that enforces that timing in two downstream branches. Such arrangements could be evolutionarily advantageous, ensuring parallel processing without interference.
3. Clock-and-Data Hypothesis
3.1 Theta Resets as Global Timing Signals
Fries (2015) and others propose that theta rhythms reset lower cortical regions during attentional shifts, organizing subsequent processing stages. EEG evidence supports this: theta activity in the PFC leads posterior cortical responses(Raposo et al., 2023).
Drawing on hippocampal analogies, theta resets in the entorhinal cortex—driven by the medial septum—enable spatial coding via phase precession (Robinson et al., 2024). A similar mechanism may operate in attention control: theta from anterior PFC resets IFJp, which gates beta (FEF) and delta (IFJa) routing.
Figure 3 summarizes oscillatory connectivity between these hubs. TMS-EEG studies show that rhythmic 6-Hz stimulation of right IFJ entrains occipital theta activity, whereas parietal stimulation does not.
This rhythmic top-down control ensures that communication happens at the right moment, allowing spatial and object streams to operate in turn without interference.
3.2 Precession and Coordination
Theta precession—a forward shift of oscillatory phase—could serve as a dynamic coordination mechanism between IFJp, FEF, and IFJa. Beta and delta bursts might align to theta phase in a multiplexed fashion, allowing direction and velocity calculations, akin to grid cell coding in navigation.
Precession could also imply that different layers of cortex process different components of a sensory event depending on theta phase, allowing even finer segregation. Such a model would predict a nested hierarchy of rhythms, with theta organizing beta and delta events in a context-sensitive manner.
This may explain how spatial and object pathways operate in lockstep yet without interference, mediated by rhythmic separation and precession.
4. Discussion and Future Directions
The “Clock-and-Data” hypothesis reframes attentional control as a form of rhythmic time-division multiplexing. A global theta clock—likely from anterior PFC—coordinates downstream beta (spatial) and delta (object) channels, gated via IFJp.
While IFJp is unlikely to originate theta activity, it plays a critical relay and gating role. Future work should use Granger causality, phase-slope index, or TMS to test directional influences.
Experimental Tests
-
TMS to IFJp: Should delay (but not abolish) phase alignment of FEF and IFJa.
-
TMS to anterior PFC: Should disrupt theta resets entirely.
-
ECoG/MEG analyses: Could reveal theta-leads-beta/delta dynamics during attentional shifts.
-
Cross-frequency coupling (CFC): Analysis between IFJp and its targets could validate the multiplexing architecture.
Theoretical and Conceptual Implications
This hypothesis also opens up conceptual links to theories of working memory and predictive coding. A rhythmic structure allows the brain to alternate between sampling external input and updating internal models. In this sense, the theta clock does more than coordinate attention; it may also align perception and prediction.
Moreover, the model suggests a unifying principle: that temporal segmentation is a general feature of cognitive control, not limited to attention. Decision-making, memory encoding, and even language comprehension might be governed by similar nested rhythms.
Challenges and Alternative Explanations
Brookshire (2022) challenges periodic models of attention, arguing that apparent rhythms may arise from aperiodic structures. However, theta coordination may still operate upstream, even if behavioral output appears irregular. A nested-rhythm model could account for both perspectives: an underlying clock drives system organization, while noisy or adaptive outputs obscure this regularity.
Furthermore, while frequency-based models provide elegant explanations, the spatial resolution of current EEG/MEG techniques limits the ability to confirm causality. It remains a challenge to dissociate overlapping sources and to prove that specific rhythms drive behavior.
5. Conclusion
This paper proposes that frequency-specific attention routing relies on a shared theta rhythm to coordinate timing between spatial (beta) and object-based (delta) streams. IFJp is positioned as a relay node, not a pacemaker, passing on theta pulses to downstream effectors. The model integrates anatomical, frequency-specific, and temporal dynamics into a coherent framework that invites both experimental and theoretical exploration.
Beyond attention, this rhythmic framework may offer a general mechanism for organizing parallel cognitive processes. By aligning signals to common clocks, the brain ensures temporal precision across distributed networks—a principle as elegant as it is powerful.
Extra
Quotes from the papers
Baldauf, D., & Desimone, R. (2014). Neural Mechanisms of Object-Based Attention. Science, 344(6182), 424–427. https://doi.org/10.1126/science.1247003
- IFJ more important for “what” is being attended than FEF:
Soyuhos, O., & Baldauf, D. (2023). Functional connectivity fingerprints of the frontal eye field and inferior frontal junction suggest spatial versus nonspatial processing in the prefrontal cortex. European Journal of Neuroscience, 57(7), 1114-1140.
 x
x
helpful starting phrases by chatty
Below are plug-and-play sentence openers organised by the same section numbers in your outline. Feel free to tweak tense, add detail, and drop in citation numbers (e.g. “[3]”) where indicated. The phrasing keeps a critical, balanced tone, nudges the reader to think beyond the obvious, and makes transitions painless.
1 Framing the Puzzle
-
“For over three decades, the field has separated attention into ‘where’ and ‘what’ streams [1]; yet this anatomical story leaves a temporal blind spot.”
-
“Why, then, do spatial and object cues seldom interfere in behaviour? One possibility is frequency-based multiplexing.”
-
“Here, I argue that oscillatory codes—rather than static pathways—provide the missing logic, a view sharpened by three provocation questions (Q1–Q3).”
2 Two Frontal Hubs
-
“Functionally, the frontal eye field (FEF) and the inferior-frontal junction (IFJ) occupy adjacent gyri but inhabit different networks.”
-
“Anatomically, FEF aligns with Glasser parcel 8Av/8C, whereas IFJ subdivides into a posterior (IFJp) and anterior (IFJa) sector [2].”
-
“Such fine-grained parcellation matters: pooling IFJp with premotor cortex can mask its ventral visual connectivity.”
3 Clock-and-Bus Hypothesis
-
“I conceptualise the system as a nested bus architecture: β rhythms set the timetable, γ rhythms carry the parcels, and slow θ/δ pulses ring the station bell.”
-
“Under Q1, one could ask whether β already suffices as both clock and carrier; a falsifiable prediction follows—β power should then covary with object identity, which it does not [4].”
-
“By contrast, a genuine two-lane highway predicts cross-frequency interactions (β phase → γ amplitude) between hubs and their visual targets.”
4 Evidence Tier 1 – Intrinsic Wiring
-
“Resting-state MEG shows the segregation before any task demands: IFJa couples to ventral cortex in γ/δ, while FEF couples to dorsal cortex in β [5].”
-
“Critically, these fingerprints survive stringent leakage control, suggesting they reflect true communication rather than common sources.”
-
“However, Smith et al. report mixed β-γ coupling for both hubs—possibly a parcellation artefact, a caveat I revisit below.”
5 Evidence Tier 2 – Task-Evoked Dynamics
5.1 Object loop
-
“When faces and houses overlap spatially, IFJ γ bursts realign FFA/PPA within ~20 ms, biasing perception toward the cued object [6].”
5.2 Spatial loop
-
“Conversely, directing covert attention to a location elevates fronto-parietal β and suppresses contralateral occipital α [7].”
5.3 Cooperation vs competition
-
“Dual-cue paradigms reveal that object and spatial loops often operate multiplicatively rather than competitively, implying genuine multiplexing.”
-
“Yet Harris et al. found only additive gains; their task, however, confounded load with cue type, highlighting a methodological pitfall.”
6 Evidence Tier 3 – Causal Manipulations
-
“Rhythmic TMS provides the cleanest test: entraining FEF at 20 Hz biases decisions conservatively, whereas 40 Hz boosts sensitivity [8].”
-
“Lesion data mirror this dissociation—cooling FEF degrades spatial gating but leaves feature filtering intact [9].”
-
“Nevertheless, we lack a study that inactivates both hubs simultaneously while tracking frequency-specific rescue.”
Text-Box 1 (Methodological Pitfalls)
-
“Source leakage: Without beamforming and pair-wise orthogonalisation, β-γ cross-talk is easily over-estimated.”
-
“Parcellation drift: The term ‘IFJ’ varies across atlases, inviting false positives.”
-
“Task-difficulty β inflation: Elevations in β can index cognitive load rather than gating.”
7 Is IFJp a θ/α Mode-Switch?
-
“IFJp exhibits slow δ/θ coupling to both dorsal and ventral streams at rest, suggesting a hub for set-shifting.”
-
“Two hypotheses diverge here: H1, IFJp generates θ pulses that alternate β and γ loops; H2, θ originates subcortically, with IFJp merely a relay.”
-
“Closed-loop θ-TMS over IFJp now allows this prediction to be tested directly.”
Text-Box 2 (Proposed Experiments)
Start each bullet with a directive:
-
“Design: Orthogonal cueing with unpredictable spatial uncertainty.”
-
“Prediction: Concurrent FEF-β and IFJ-γ coherence will forecast dual-cue performance.”
8 Synthesis Across Tiers
-
“Across rest, task, and perturbation studies, a convergent pattern emerges: β establishes the gate, γ delivers the payload, and θ schedules the swap.”
-
“The two strongest counter-studies remind us that oscillatory roles are not immutably hard-wired but context-dependent.”
Figure 2 Legend (opening lines)
-
“Panel A depicts the nested architecture: a θ/δ master clock (grey) envelops a β gate (blue) and γ payload bursts (red).”
-
“Panel B aligns behavioural RT with the phase of these oscillations, illustrating the functional read-out.”
9 Outlook for the Thesis
-
“Chapter 2 will operationalise these hypotheses in a preregistered MEG study.”
-
“By integrating oscillatory multiplexing with behavioural indices, the thesis aims to refine how cognitive control is quantified in humans.”
Transitional Phrases & Contrast Cues
-
“Critically, however…” | “In seeming contradiction…”
-
“At first glance this supports X; a closer inspection suggests Y.”
-
“This finding dovetails with…” | “The picture blurs when…”
Use these starters to leap past writer’s block, drop in your numbers, and keep the prose crisp yet skeptical. Happy drafting!
see also
Tags: neuroscience science
Superlink: 050 🧠Neuroscience
Outline Seminar Paper Brain Connectivity
Project Proposal Seminar Brain Connectivity
Source
Created: 05-06-25 18:34