Tutorial Script – Lecture 1: Intro & Spinal Cord
Date: Wednesday, 15 April 2026
Duration: 90 minutes
Lecture slides: AC_MS_1_SpinalCord_new.pdf
0. Before the Tutorial (prep, not spoken)
Expected student questions:
- “What exactly is the difference between upper and lower motor neurons?”
- “Is the stretch reflex the same as the knee-jerk reflex?”
- “What does feedback vs. feed-forward mean in practice — can you give me an example?”
- “Why can’t the spinal cord regenerate? I thought neurons can grow back.”
- “What are force fields and why are they in Lecture 1 if they’re about frogs?” (Force fields appear in the course index under Lecture 2 but in the exam questions for Lecture 1 — clarify: force fields are a Lecture 1/2 boundary topic. Check which exam question list students are looking at.)
- “Do CPGs exist in humans, or only in animals?”
Common misconceptions to watch for:
- Confusing UMN and LMN lesion symptoms (students often mix up: flaccid = LMN, spastic = UMN)
- Thinking feedback control is always better than feed-forward — neither is universally better; they complement each other
- Believing reflexes are completely hard-wired and invariant — lecture explicitly shows they are context-dependent and learnable
- Confusing the corticospinal tract crossing (UMN decussates → contralateral control) with LMN staying ipsilateral to the muscle it controls
- Thinking the PD controller analogy means the brain is literally computing a derivative — it’s a functional analogy for stiffness + damping implemented by the stretch reflex
Personal prep notes:
- Know the PD controller formula: Q(t) = Pθ(t−τ) + Dθ̇(t−τ) — P = stiffness term, D = damping term
- Force field experiment: Mussa-Ivaldi lab, frog spinal cord, linear superposition of force vectors — this is Lecture 1 content in the slides even if it bridges to Lecture 2 notes
- Review Nogo protein story: Schwab (inhibitory CNS myelin) vs. Zheng et al. 2005 (NgR mutants didn’t show enhanced regeneration) — the lecture explicitly acknowledges the controversy
- CPG half-center model: mutual inhibition of two oscillatory pools → alternating limb movement; “GO” signal from brain is tonic, not alternating
Fact-check note from vault:
ℹ️ The vault note on Spinal cord.md says “Descending fibers connect the motor cortex to the spinal cord in an ipsilateral fashion” — this is incorrect. The corticospinal tract crosses the midline (decussates at the medullary pyramids), giving contralateral control. The vault note should not be trusted on this point. The lecture slides are correct.
1. Opening & Hook (5 min)
You’re walking along a cobblestone street at night. Your foot hits an unexpected dip. Before you even register what happened consciously, you’ve corrected your balance. How?
If the brain had to send a signal all the way down to your foot, process it, and send a correction back up — the signal alone takes roughly 80–100 ms each way just for nerve conduction, not counting cortical processing. At a normal walking speed, you’d be on the ground before the cortex could help.
Yet we don’t fall. Something else is doing the work.
Opening question for the group:
“If the brain can’t react fast enough to catch us when we stumble, what is actually keeping us upright — and what does that tell us about how the nervous system is organized?”
(Goal: surface the key idea of computational offloading. Let students guess. Draw out: local circuits, reflexes, physical properties of muscles. Then connect this to the lecture’s central argument: the spinal cord is an “edge computing” device.)
2. Link to Last Week (5 min)
This is the first tutorial — no previous week to link to.
Use this time instead to establish the course-wide frame:
König’s central argument throughout this course: the classic “perception → decision → action” sequence is a caricature. These systems are highly parallel and integrated. We’re starting from the bottom — the spinal cord — because understanding what the spinal cord handles autonomously tells us what the cortex doesn’t need to do.
Ask students:
“Before this course, what did you think motor control was? Cortex sends command, muscles execute — is that the whole story?”
(Usually students have a top-down, cortex-centric mental model. This session should begin dismantling it.)
Anchor this in everyday terms: walking, typing, balancing — none of these require conscious micro-management. The challenge of this lecture is asking why not, and what exactly is doing the work instead.
3. Motor System Overview: The Big Picture (15 min)
Core Message
The motor system is a hierarchy of control loops, not a single command structure. Each level handles a different timescale and specificity of control. The cortex sets goals; subcortical structures and the spinal cord implement them.
Explanation Approach
Draw the hierarchy on the board (students should be able to reproduce this — it’s a direct exam question):
Perception → Decision → Action
|
┌────────────┴────────────┐
Cortical Areas Cerebellum
(Planning, M1) (Timing, Precision)
| |
Basal Ganglia ───────── Brainstem
(Action selection) (Posture, goal-directed)
|
Spinal Cord
(Reflexes, CPGs, Rhythm)
|
Muscles
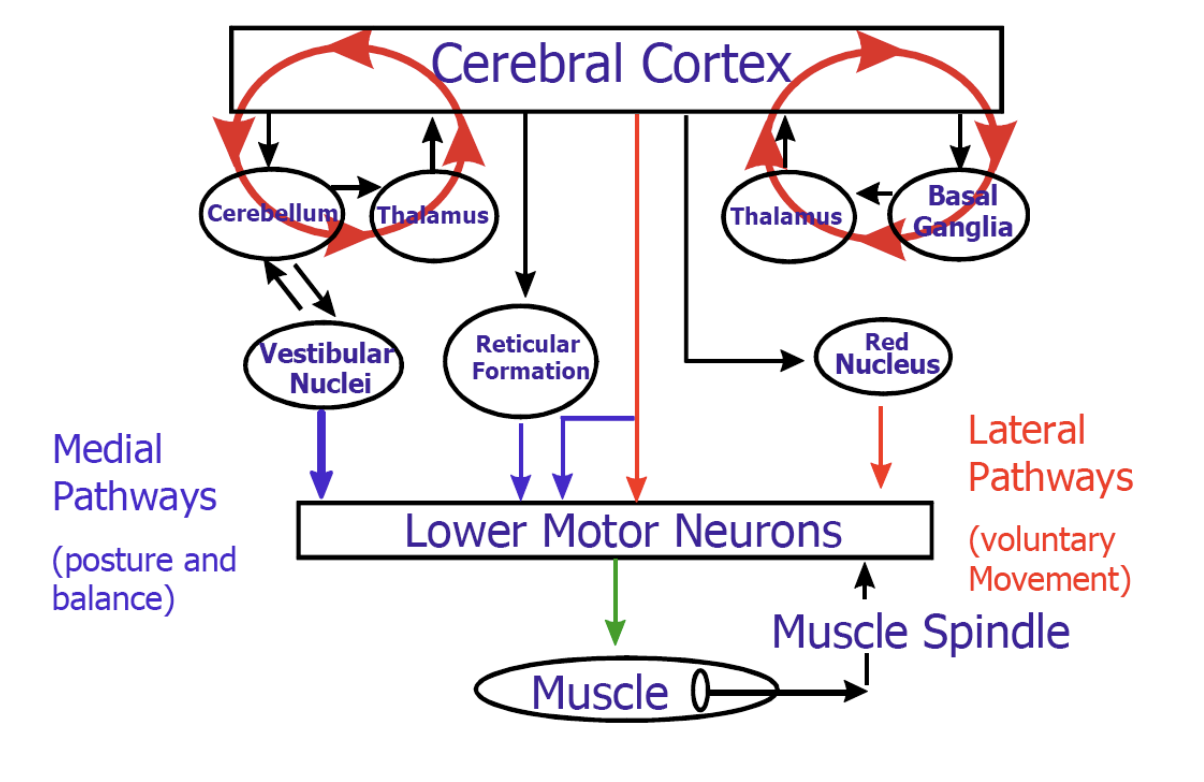
Key point: Two types of motor neurons form the final common pathway:
- Upper Motor Neurons (UMN): cell bodies in CNS (cortex, vestibular nuclei, reticular formation, red nucleus); axons project down to LMNs
- Lower Motor Neurons (LMN): cell bodies in brainstem or spinal cord; axons project out to striated muscle — the final common pathway, the bottleneck
Discussion Questions
- “Why does it make sense architecturally to have the LMN as the ‘final common pathway’? What does this buy you computationally?”
- “The cerebellum and basal ganglia both influence movement, but neither projects directly to muscles. What does that tell us about their role?”
- “In the perception → decision → action scheme: where does the stretch reflex fit? Is it perception, decision, or action — or none of the above?”
Common Misconceptions
- “The cortex controls everything” → The cortex sets high-level goals; the spinal cord, cerebellum, and BG handle enormous amounts of computation independently
- “UMN = in the brain” → UMN cell bodies are in multiple CNS locations, not only cortex (also vestibular nuclei, red nucleus, reticular formation)
Exam Relevance
- Exam Q1: “Draw a sketch of the modules of the overall mammalian motor system and their interactions.” — students need to produce a labeled diagram. Practice this now.
- Good answer: labels Cortex, Cerebellum, BG, Brainstem, Spinal Cord with directional arrows and functional labels; includes UMN/LMN distinction
4. Feed-Forward vs. Feedback Control (15 min)
Core Message
The motor system uses both strategies. Feedback is flexible but slow and can oscillate. Feed-forward is fast but requires an accurate internal world model. The spinal cord implements a hybrid via the stretch reflex.
Explanation Approach
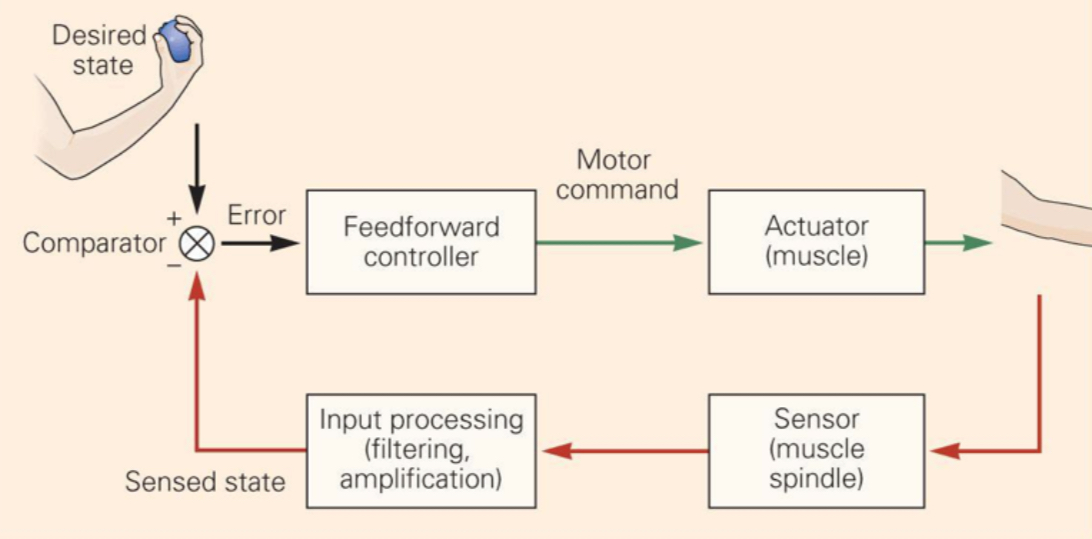
Feedback (closed-loop):
- Compare actual state to desired state
- Correct errors in real time
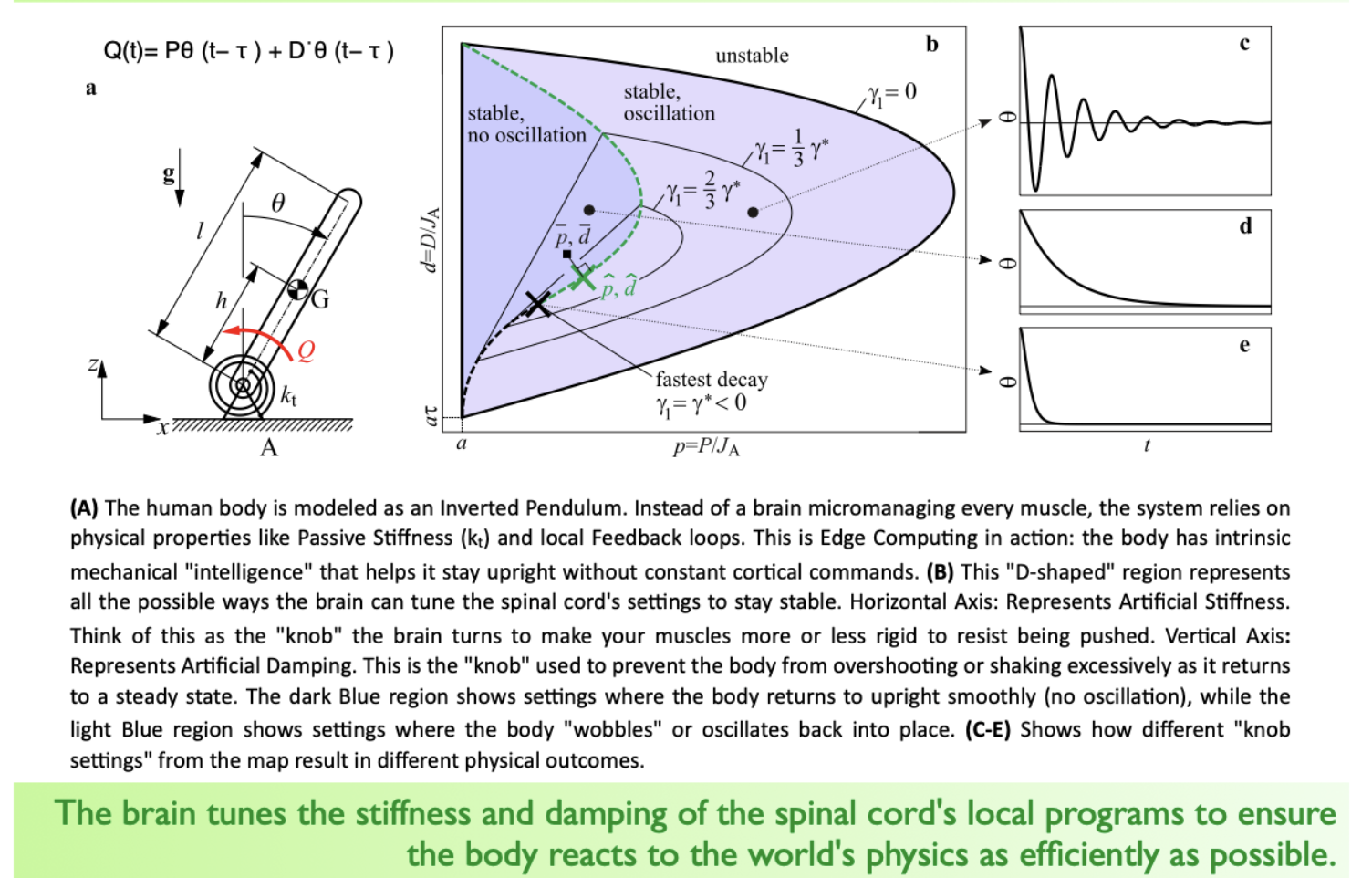
- Problem: delay τ between sensing and acting → if τ is large, oscillations emerge (stick-balancing demo is the lecture’s example)
- Formula from lecture: Q(t) = Pθ(t−τ) + Dθ̇(t−τ) — this is a PD controller with delay
- The D-shaped stability region in the lecture graph shows: not all P/D settings work; some lead to oscillation, some to smooth settling
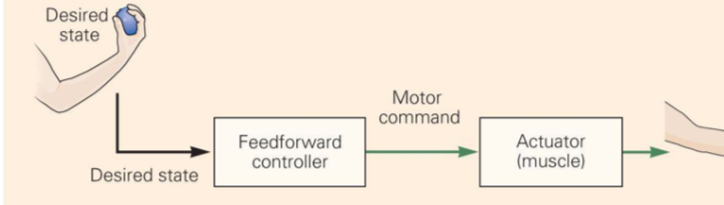
Feed-forward (open-loop):
- Uses a prediction / internal model to act before sensing error
- Fast — saccades are the classic example (too fast for visual feedback to correct mid-flight)
- Error-prone: if the model is wrong, the error is not caught
Analogy: Driving with vs. without GPS rerouting:
- Feedback = GPS that constantly recalculates based on where you actually are
- Feed-forward = driving from memory — fast, but if you’ve memorized the wrong route, you end up in the wrong place
Discussion Questions
- “Think of a skilled movement you do automatically — say, typing or catching a ball. Which parts of that movement are feed-forward, and which are feedback-driven?”
- “If feedback control can lead to oscillations, why do we use it at all? What would a purely feed-forward motor system fail at?”
- “The lecture mentions the spinal cord as an ‘edge device’ like in IoT computing. What does offloading computation to the spinal cord buy the brain — and what does it cost?”
Common Misconceptions
- “Feedback is always better because it corrects errors” → Feed-forward is necessary for fast movements where feedback is too slow to be useful; real systems combine both
- “The stretch reflex is pure feedback” → It is a local feedback loop, but the brain modulates its gain via corticospinal input — so it’s a tunable feedback circuit, not a fixed one
Exam Relevance
- Exam Q2: “Explain and contrast the concepts of feed-forward and feed-back control.”
- Good answer must include: definition of each, speed/delay tradeoff, error correction capability, oscillation risk in feedback, world-model requirement for feed-forward, concrete examples for each
5. The Stretch Reflex: The Spinal Cord as a PD Controller (15 min)
Core Message
The stretch reflex is a monosynaptic, hardwired feedback loop that implements stiffness and damping at the hardware level — this is the biological PD controller that keeps us upright without cortical involvement.
Explanation Approach
Walk through the circuit step by step:
- Muscle is stretched → muscle spindle (intrafusal fibers) detects length and velocity changes
- Ia afferent (fastest myelinated fiber) carries signal via dorsal horn → alpha motor neuron (ventral horn) — monosynaptic!
- Alpha motor neuron activates the same muscle (and synergists) → resists the stretch
- Reciprocal inhibition: Ia afferent also activates an inhibitory interneuron → inhibits alpha motor neuron of antagonist → antagonist relaxes (otherwise co-contraction would lock the joint)
- Ib afferents (Golgi tendon organs): sense force (not length); at very high forces, they inhibit the alpha motor neuron → prevents tendon damage (autogenic inhibition)
The PD analogy:
- P (proportional) = responds to the amount of stretch (position error)
- D (derivative) = responds to the rate of stretch (velocity)
- Together: the muscle stiffness (P) and damping (D) are implemented by the Ia afferent’s combined sensitivity to length and velocity
Corticospinal modulation: The cortex doesn’t bypass this reflex — it tunes it. The gain of the stretch reflex can be modulated by descending input, which is how the cortex plays “on the keyboard of the spinal cord” (König’s phrase).
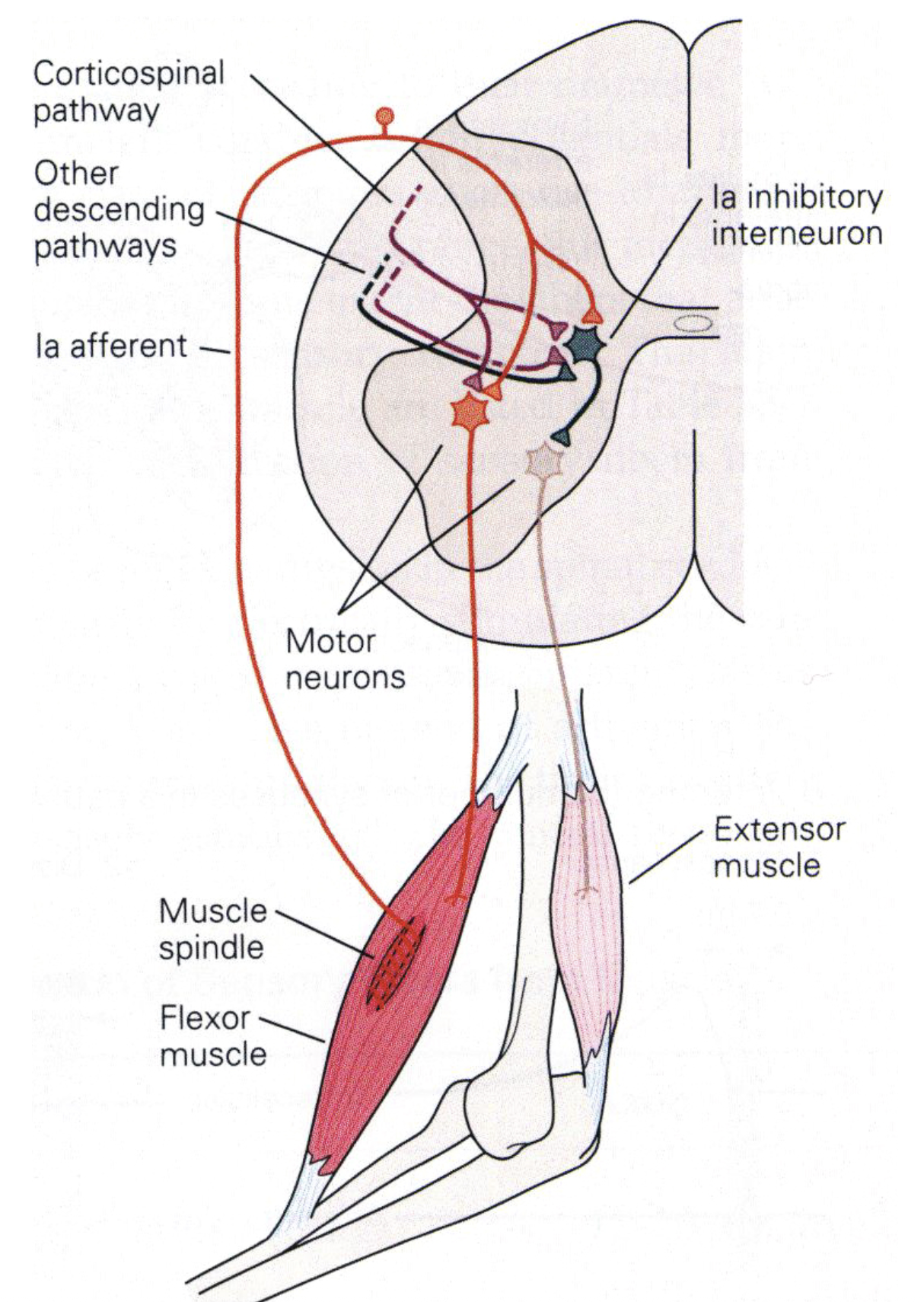
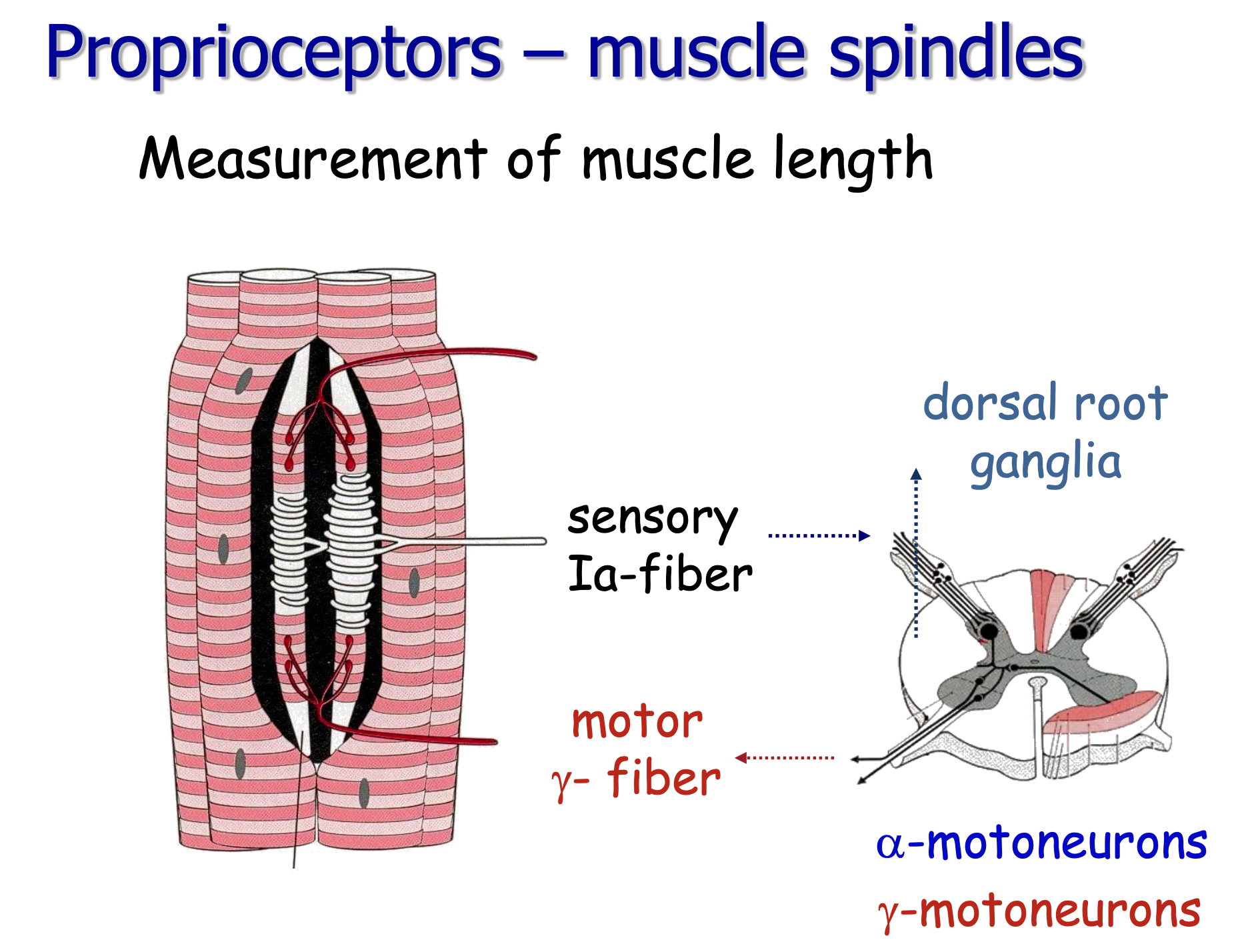
Discussion Questions
- “The stretch reflex is monosynaptic — why is that architecturally significant? What does having only one synapse in the loop buy you?”
- “Reciprocal inhibition means activating the bicep automatically inhibits the tricep. How does this simplify the commands the brain needs to send?”
- “The lecture says ‘the spinal cord is performing calculus at the hardware level.’ What calculus is it performing, and how does a biological circuit do that?”
Common Misconceptions
- “The Ia afferent connects directly to the muscle” → Ia afferent connects to the alpha motor neuron (in the ventral horn), which then projects to the muscle. The reflex arc is: spindle → Ia afferent → alpha MN → muscle.
- “Ib afferents = same as Ia” → Ia = spindle (length/velocity), Ib = Golgi tendon organ (force). Different signals, different functions.
- “Reciprocal inhibition means the antagonist is completely silent” → Its tone is reduced, not abolished. The level of inhibition is graded.
Exam Relevance
- Exam Q5: “What are the mechanisms underlying the stretch reflex?”
- Good answer: Ia afferent → alpha motor neuron (monosynaptic); reciprocal inhibition via interneuron; Ib afferent / Golgi tendon organ for force sensing; mention corticospinal modulation of reflex gain
6. Central Pattern Generators (10 min)
Core Message
For rhythmic movements like walking, the spinal cord doesn’t need the brain to send alternating “left, right, left, right” commands. A half-center oscillator circuit generates the rhythm autonomously. The brain sends a tonic “GO” signal.
Explanation Approach
The Graham Brown experiment (mentioned in lecture): decerebrate cat on a treadmill. Cortex is severed; spinal cord receives only tonic electrical stimulation. Cat walks, trots, gallops — transitioning automatically as treadmill speeds up. The spinal cord is generating the entire locomotor pattern.
The half-center model:
- Two pools of neurons mutually inhibit each other
- Only one can be “on” at a time
- When one fatigues or is overridden by sensory input, it releases the other
- Result: alternating activation = stepping pattern
The key insight: This is embodied cognition — the physics of the body (inertia, contact forces detected by stretch receptors) participates in the computation. The CPG doesn’t just drive muscles; it’s coupled to the body’s mechanics.
Two inputs to CPG:
- Tonic descending drive from brain (sets “GO” state and speed)
- Afferent sensory feedback (modulates timing based on actual ground contact, limb loading)
Discussion Questions
- “If CPGs can generate locomotion without the brain, what does the brain actually need to do to control walking?”
- “CPGs were demonstrated in decerebrate cats. How would you test whether CPGs exist in humans, given ethical constraints?”
- “The CPG model makes locomotion more robust to perturbations. Why? What does having local feedback loops buy you compared to pure central control?”
Common Misconceptions
- “CPGs are only in spinal cord animals like cats or frogs” → Evidence for CPG-like circuits in humans exists (humans with complete spinal cord injury can show stepping-like activity under certain stimulation conditions)
- “CPGs eliminate the need for sensory feedback” → CPGs can operate without feedback (fictive locomotion), but normal locomotion relies on sensory feedback to adapt to terrain
7. UMN vs. LMN Lesions (10 min)
Core Message
The location of a lesion determines the syndrome. LMN lesion = flaccid paralysis + areflexia + atrophy. UMN lesion = spastic paralysis + exaggerated reflexes + Babinski sign. The difference reveals the functional anatomy of the motor system.
Explanation Approach
Draw a simple table on the board:
| Feature | LMN Lesion | UMN Lesion |
|---|---|---|
| Tone | Flaccid (hypotonia) | Spastic (hypertonia) |
| Reflexes | Absent (areflexia) | Exaggerated |
| Atrophy | Yes (denervation) | No (or minimal) |
| Babinski | Absent | Present |
| Paralysis | Yes (all excitation lost) | Yes (initially flaccid → spastic) |
Why is UMN lesion eventually spastic? The descending cortical tract normally inhibits some spinal reflex circuits. When that inhibition is removed, the stretch reflexes become hyperactive → spasticity. “Spinal shock” in acute UMN lesions = temporary loss of all spinal reflex activity before spinal circuits become hyperreactive.
Babinski sign: Stroking the sole → toes dorsiflex and fan out (plantar extension). In normal adults: plantar flexion (curling inward). The Babinski sign = loss of cortical inhibition of the spinal withdrawal-related circuit. It’s the spinal cord reverting to a “default” state without cortical override.
Hemiplegia anatomy: UMN lesion above the decussation (in internal capsule, above medullary pyramids) → contralateral hemiplegia. UMN lesion below the decussation → ipsilateral weakness (rare, mostly cervical cord).
Discussion Questions
- “Why does denervation atrophy happen in LMN lesions but not in UMN lesions? What does this tell us about the trophic relationship between motor neurons and muscle?”
- “The Babinski sign is present in healthy infants until about age 1-2. What does that tell us about cortical development and its relationship to spinal cord circuits?”
- “A patient presents with spastic paralysis and exaggerated knee-jerk but no Babinski sign. Where might the lesion be — and why might Babinski be absent?”
Common Misconceptions
- “Flaccid = no movement, spastic = more movement” → Both produce paralysis. The difference is in tone and reflex excitability, not in the patient’s ability to move voluntarily (both are impaired or absent)
- “UMN lesion only affects the cortex” → UMN can be anywhere from cortex to the synaptic contact with the LMN in the spinal cord. Brainstem lesions can produce UMN signs.
- “Babinski is a pathological reflex unique to UMN lesions” → More precisely: it’s the spinal cord’s default response when cortical inhibition is absent. It’s “physiological” in infants and becomes pathological only once myelination and cortical maturation are complete.
Exam Relevance
- Exam Q3 & Q4: “What is happening upon a lesion of the upper and lower motor neurons?” / “What is happening upon a lesion of the upper motor neurons?”
- These are two separate exam questions — students need to know both syndromes distinctly, not just “UMN vs LMN”
- Good answer for UMN: initial spinal shock → spasticity, exaggerated tendon reflexes, Babinski, hemiplegia if unilateral, contralateral if above decussation
8. Spinal Cord Lesions & Recovery (5 min)
Core Message
Spinal cord injury is devastating and currently irreversible, primarily because CNS axons cannot regenerate — unlike PNS axons. The Nogo protein story illustrates how even promising findings in neuroscience can fail to translate.
Key Points
- 12,000 new cases/year in the US; ~273,000 living with SCI; nearly half aged 16–30
- CNS (including spinal cord) ≠ PNS: no axon regeneration in CNS; PNS axons can regrow ~1 mm/day
- Nogo protein on CNS myelin inhibits axon regrowth → idea: inhibit the inhibitor (Schwab’s group)
- But: Zheng et al. 2005 showed NgR-deficient mice did NOT show enhanced corticospinal regeneration → the simple Nogo story is likely incomplete
- Recent treatment directions: epidural stimulation allowing voluntary movement (“considering the movement might help therapy” — the lecture’s final slide)
ℹ️ Note: The Nogo/NgR story is an excellent example of how exciting initial findings can face replication and mechanistic challenges. Good to discuss with students how science self-corrects.
Discussion Question
“The lecture ends with ‘considering the movement might help therapy.’ What does this mean — and how does it connect to everything we covered about CPGs and spinal plasticity?”
Exam Relevance
- Exam Q6: “Which mechanisms limit recovery after spinal cord lesions?”
- Good answer: CNS lacks PNS-like axon regeneration; surgical inaccessibility; Nogo/inhibitory myelin proteins; incomplete understanding of circuit reorganization
9. Exam Question Round (15 min)
Work through these verbatim — ask students to answer first, then correct/extend.
Question 1: “Draw a sketch of the modules of the overall mammalian motor system and their interactions.”
Expected key points:
- Hierarchical diagram: Cortex, Cerebellum, Basal Ganglia, Brainstem, Spinal Cord, Muscles
- Direction of arrows (descending motor, ascending sensory feedback loops)
- UMN/LMN distinction labeled
- Mention that perception and action are integrated (not strictly serial)
Typical mistakes:
- Missing Brainstem as a separate level
- Forgetting to label which structures are CNS (UMN) vs. the final LMN pathway
- Drawing a linear chain instead of a parallel hierarchy with feedback loops
Question 2: “Explain and contrast the concepts of feed-forward and feed-back control.”
Expected key points:
- Feedback: compares actual to desired state, error correction, slow, risk of oscillation with delay
- Feed-forward: uses internal world model, no sensory correction, fast, error-prone
- Both are used in the real motor system simultaneously
- Concrete example each: e.g., catching a thrown ball (feed-forward dominant) vs. slowly adjusting grip force on a fragile object (feedback dominant)
Typical mistakes:
- Saying “feed-forward is better” or “feedback is better” without acknowledging the tradeoff
- Not mentioning the delay/oscillation problem in feedback
- Forgetting that feed-forward requires a world model
Question 3: “What is happening upon a lesion of the upper and lower motor neurons?”
Expected key points:
- LMN: flaccid paralysis, areflexia, muscle atrophy (denervation), fasciculations possible
- UMN: initially flaccid (spinal shock) → spastic paralysis, exaggerated tendon reflexes, Babinski sign, hemiplegia contralateral to lesion if above decussation
- UMN lesion leaves the reflex arc intact (spinal cord still connected to muscle) but removes descending modulation
Typical mistakes:
- Confusing which syndrome is which (students mix flaccid/spastic)
- Forgetting spinal shock as the acute UMN presentation
- Not mentioning Babinski
- Not explaining why reflexes are exaggerated in UMN lesion (disinhibition)
Closing (3 min)
Take-Home Message:
“The spinal cord is not a simple cable connecting brain to muscle — it is a sophisticated computing device that handles reflexes, rhythms, and muscle coordination autonomously, freeing the cortex to focus on goals rather than mechanics. The brain doesn’t micromanage muscles; it plays on the keyboard of the spinal cord.”
Preview of next week: Lecture 2 – Cerebellum. If the spinal cord handles fast local reflexes, what does the cerebellum add? Answer: timing and precision — especially for learned, predictive movements. We’ll see how the cerebellum functions as a feed-forward controller and how classical conditioning illuminates cerebellar learning.
Appendix: Images Available in Vault
- — Motor system overview diagram
- — Stretch reflex circuit diagram
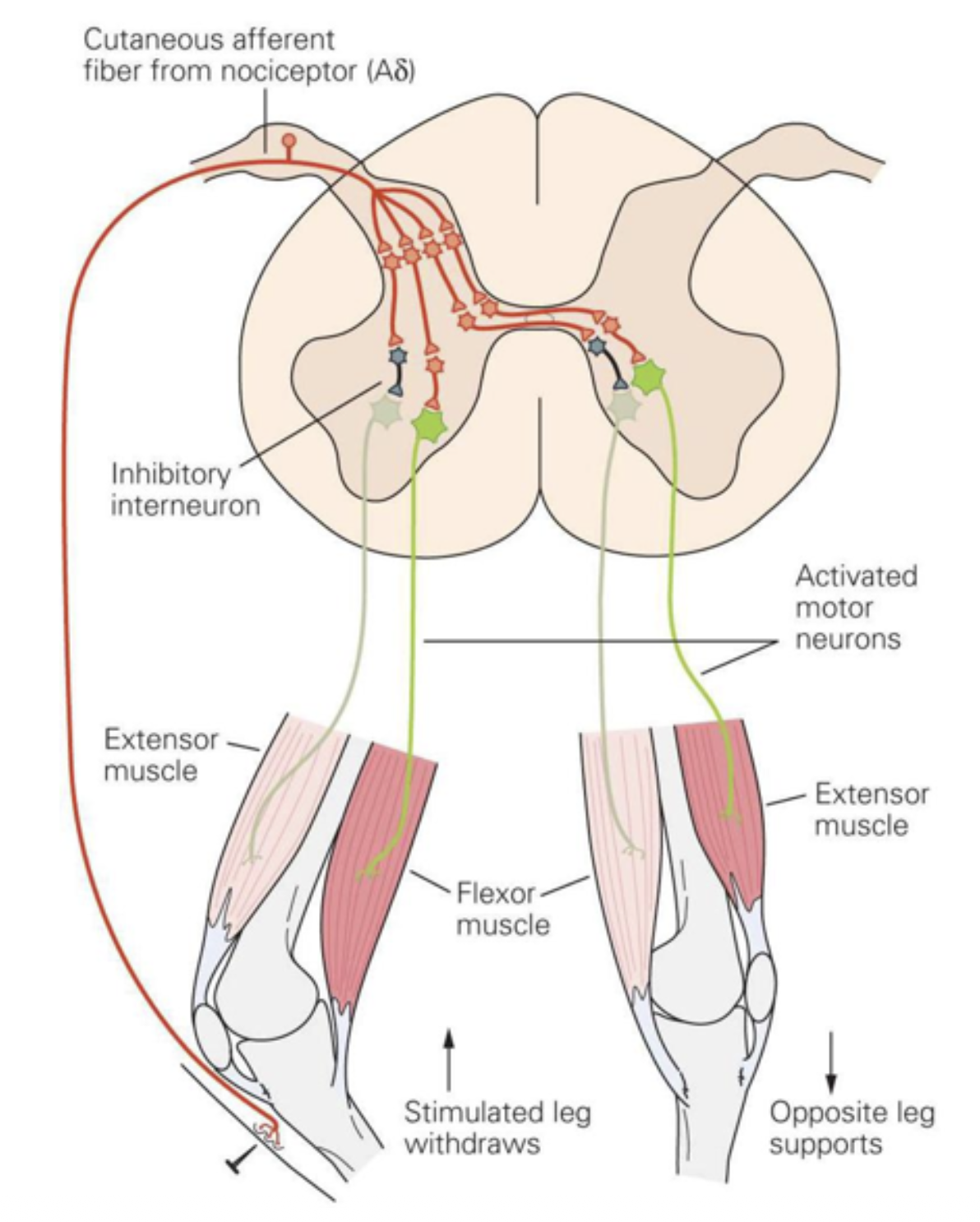 — Spinal cord anatomy
— Spinal cord anatomy- — Muscle spindle diagram
- — Offloading challenge / edge computing analogy
- — Feed-forward control diagram
- — Feedback control diagram
Tags: motorsystem tutor