Tutorial Script – Lecture 4: Motor Cortex
Date: Wednesday, 7 May 2026
Duration: 90 minutes
Lecture slides: AC_MS_4_MotorCortex.pdf
0. Before the Tutorial (prep, not spoken)
Likely questions from students:
- “What exactly is the population vector and how do you calculate it?”
- “What’s the difference between SMA and premotor cortex — they’re both BA6?”
- “Why does M1 have a smaller Layer 4 than somatosensory cortex?”
- “What does repetition suppression actually measure — is it just about repeated images?”
- “DBS stimulates but you said it inhibits — how can stimulation cause inhibition?”
- “Birbaumer’s BCI is EEG-based — why is EEG different from the BrainGate implant?”
Known misconceptions to actively address:
- Conflating SMA and premotor cortex (both BA6, but different locations and functions)
- Thinking the homunculus is anatomically accurate (it’s representationally weighted, not proportional)
- Believing M1 and S1 are cleanly separated at the central sulcus — they are “more of a bias than absolute”
- Confusing “preferred direction” with “only active direction” — neurons have cosine tuning curves, not binary responses
- Thinking “imagined = same as executed” at the neural level — it’s not (SMA only, no M1/S1)
- Assuming BrainGate reads thoughts, not movement directions
Things to look up again before tutorial:
- Georgopoulos 1986 (original PV paper), Schwartz et al. 2007 J Physiol (motor illusion)
- Grafton & de Hamilton 2007 Hum Mov Sci (repetition suppression, distributed contributions)
- Birbaumer et al. 1999 Nature (first BCI, 0.5 bit/s, locked-in patient)
- Graziano & Aflalo 2007 Neuron (8 competing topographic proposals, 3-dimension model)
- Corticospinal tract numbers: M1 40%, PM 30%, S1 20%; ~1M fibres; 90% cross
1. Opening & Hook (5 min)
“You’re watching someone pick up a glass. You don’t move. But something in your motor cortex fires anyway — neurons that would fire if YOU were picking up the glass. Now imagine the reverse: your hand moves in a direction you didn’t intend, but your brain thinks it moved correctly. Which part of the motor cortex holds the real ground truth?”
Let the group react. This sets up the whole lecture’s tension: the motor cortex is not a simple command center but a distributed, hierarchical system where what you plan (PMv) and what you actually execute (M1) can come apart.
Alternative hook (if group is quiet):
“In 1999, a completely paralyzed man typed a letter to his doctor using only his brain waves and a computer. No muscles. No implants. Just EEG. At 0.5 bits per second. That was the first brain-computer interface. What has happened since — and what does the motor cortex have to do with it?”
(Sets up the BCI section at the end of the tutorial — creates a narrative arc.)
2. Link to Last Week (5 min)
Last week (Lecture 3): Basal Ganglia = the selection gate. The BG answer which movement gets through via tonic inhibition of the thalamus that is selectively released. The output goes: GPi → Thalamus (VA/VL) → Motor Cortex.
This week: the motor cortex — how the movement is actually specified and commanded. M1 is where the BG’s “go” signal lands, and where it gets translated into a directional command that travels down to the spinal cord.
Bridge question:
“If the BG decide which action gets permitted, and the thalamus relays that decision to motor cortex — what is motor cortex actually doing? Is it just passing the signal along, or computing something new?”
(Guide toward: M1 is computing the movement direction itself — direction tuning, population coding — which is quite different from the selection gating the BG do.)
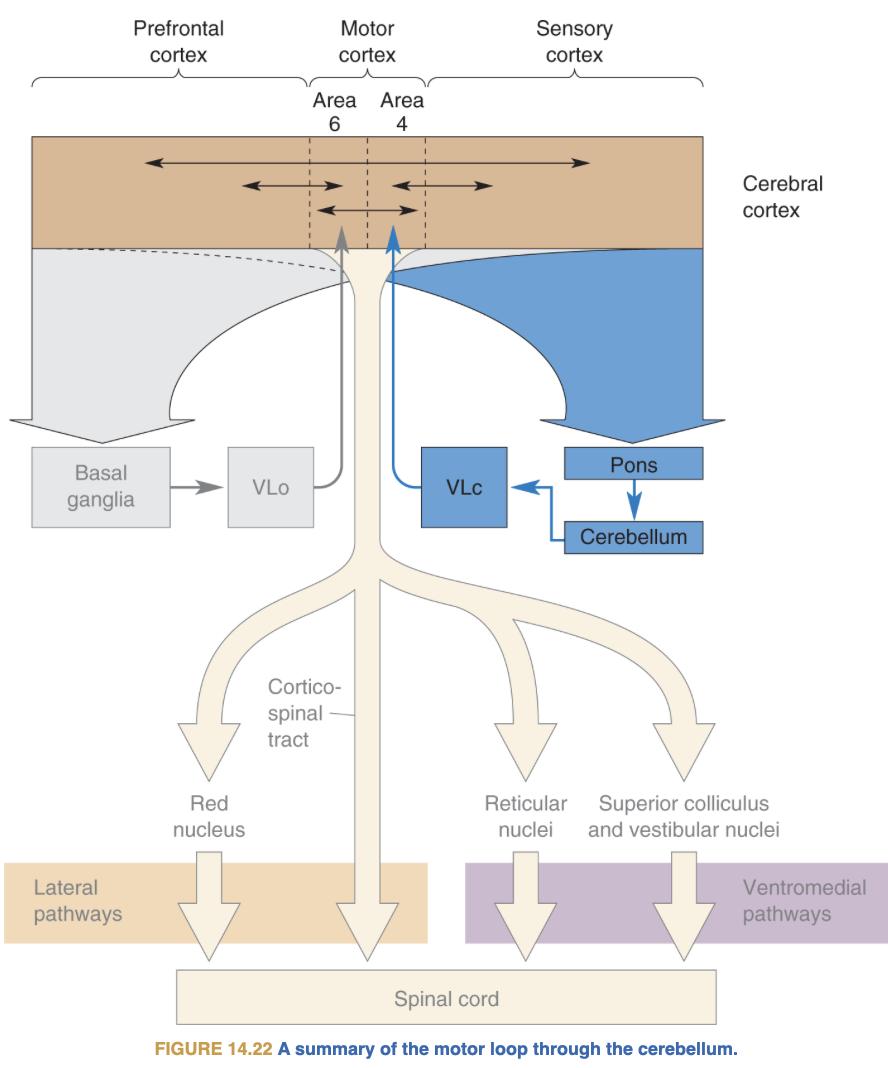
3. Four Cortical Motor Areas (12 min)
Core Message
The motor cortex is not a single area. Four distinct cortical regions form a hierarchy — from abstract intention to concrete muscular command.
The Four Areas
| Area | BA | Location | Key Function | Lesion Effect |
|---|---|---|---|---|
| M1 | 4 | Precentral gyrus | Executes movement; corticospinal output | Contralateral paralysis |
| SMA | 6 (medial) | Medial wall | Self-initiated movement, preparation, procedural memory | Mutism, akinesia; speech returns but non-spontaneous |
| Premotor (PM) | 6 (lateral) | Lateral convexity | Externally-guided movement; grasp planning; F2/F4/F5/F7 | Loss of externally cued movement coordination |
| FEF | 6+8 | Caudal middle frontal gyrus | Voluntary saccades, spatial attention, spatial working memory | Contralateral gaze deviation |
Also note: Broca’s (BA44/45), Cingulate motor cortex, Orbito-prefrontal cortex.
Cytoarchitectonic rule: Sensory cortices have a prominent Layer 4 (receives thalamic input). Motor cortex is agranular — Layer 4 is small or absent. Motor cortex is primarily an output structure.
Discussion Questions
-
“SMA lesions cause not just movement poverty (akinesia) but mutism. Why would a ‘motor area’ affect speech — and why does speech return spontaneously but non-spontaneously?”
(Guide: SMA is involved in self-initiated, internally driven movement — including internally driven speech. Lesion removes self-initiated output; speech returns through other routes.) -
“Both SMA and Premotor cortex are called ‘Brodmann Area 6.’ Why is that an unhelpful label — and what anatomical landmark would you use instead?”
(Guide: BA6 covers a vast swath; SMA is on the medial wall, PMC is on the lateral convexity — they’re anatomically and functionally distinct despite the same BA label.) -
“The premotor cortex projects to M1 but also partly directly to the spinal cord. If premotor has direct spinal output, how is it different from M1?”
(Guide: PM’s spinal projections are sparser and less specific; PM is still primarily a planning area; M1 has the main corticospinal output and Betz cells.)
Common Misconceptions
- “SMA = premotor” → Both BA6 but anatomically separated; SMA is internally driven, PMC is externally driven
- “FEF is purely eye movement” → Also: spatial attention, spatial working memory, evolving decision encoding
4. Activation Patterns — Who Does What? (8 min)
Core Message
The recruitment of motor areas depends critically on the complexity and learning stage of the movement. This gives us a window into the functional division of labor.
The Four Conditions (draw on board or verbal walkthrough)
Simple movement (one finger flexion):
- Active: M1 + S1 — both co-active even for the simplest voluntary movement
- Note: even “simple” movement always involves somatosensory feedback
Complex movement sequence (1-3-2-1-3-2 with fingers):
- Active: SMA + M1 + S1 + Inferior Prefrontal Cortex
- Key: primary areas (M1, S1) are lateralized (contralateral hemisphere only)
- Higher motor areas (SMA, premotor, prefrontal) are bilateral — both hemispheres engaged
Imagined movement (same sequence, not executed):
- Active: SMA only (bilateral) — no M1, no S1
- “Imagined movements remain limited to SMA”
Skilled (old) vs. New movements:
- New > Old (red — learning): SMA, Premotor, Parietal, Lateral Prefrontal, Hippocampus
- Old > New (blue — execution): Cerebellum — more active for well-learned automatized movements
- “The role of motor areas differs in learning and execution of movements”
Discussion Questions
-
“If imagined movements only activate SMA but not M1, could you use imagined movements as a BCI input? What would be the advantage — and the problem?”
(Guide: advantage = no movement needed; problem = SMA signal is vaguer/harder to decode than M1 directional signals; also no somatosensory feedback loop for calibration.) -
“Why would the cerebellum be MORE active for a skilled, well-learned movement than for a new one? Isn’t learning the cerebellum’s job?”
(Guide: Cerebellum = supervised learning machine = prediction + error correction. For a well-executed sequence, it needs to run the prediction model at full speed. For new movements, it hasn’t built the model yet — it’s recruiting cortical areas to consciously sequence. Habitual execution = maximal cerebellar forward model use.) -
“Complex movement activates inferior prefrontal cortex bilaterally. What computational role might prefrontal cortex be playing during a finger sequence — especially on the non-dominant (ipsilateral) side?”
(Guide: working memory for the sequence order; attention allocation; response suppression of the non-moving hand — all bilateral processes.)
5. Homunculus, M1 Topography & Graziano’s Model (12 min)
Core Message
The body is mapped onto motor cortex — but the map is distorted, fuzzy, and contested. The classic homunculus is a useful first approximation that breaks down at the microscopic level.
The Homunculus
- Penfield & Boldrey (1937/1950s): Electrical stimulation of exposed M1 in awake neurosurgical patients → mapped movement representations
- Distortion reflects motor complexity, not body size: hands and lips are enormous; trunk is tiny
- Somatotopy: face = lateral, feet = medial; contralateral control
- Two maps: M1 (motor) and S1 (somatosensory) — largely congruent but “the division is more of a bias than absolute”
- S1 has sites on the pre-central gyrus; M1 has sites on the post-central gyrus
- Microscopic scale: movement of a given body part (e.g., digits) evoked from multiple foci — “on a microscopic scale the map is NOT ordered”
Discussion Questions
-
“If you practice a complex piano piece for 3 months, what would you expect to happen to the hand representation in M1 — and what mechanism would drive that change?”
(Guide: expansion of hand area, invasion of neighboring representations — Merzenich experiments. Mechanism: activity-dependent plasticity, LTP at active cortico-cortical synapses.) -
“Graziano & Aflalo (2007) proposed 8 different competing models for how M1 is organized topographically. Their own model uses three competing dimensions. Why would optimizing one dimension (e.g., somatotopy) ‘scramble’ the others (ethological action category, spatial hand position)?”
(Guide: the 2D cortical sheet is finite. If you sort neurons by body part, you can’t simultaneously sort them by action type AND hand location — these are independent axes in a multi-dimensional space being flattened onto 2D. The solution is a compromise — “like attracts like” locally, none perfectly globally.) -
“The Graziano model predicts ‘ethological action categories’ as zones (reach-to-grasp, defense, hand-to-mouth, etc.). What does the existence of these action-category zones tell you about what the motor cortex is actually encoding — body part movements or goal-directed actions?”
(Guide: suggests M1 is not purely a “muscle controller” but is organized around ecologically meaningful action patterns — supports a more hierarchical view where M1 combines movement and goal.)
Common Misconceptions
- “The homunculus map is fixed and precise” → it’s plastic and fuzzy; overlapping representations; modified by experience
- “M1 and S1 are cleanly separated at the central sulcus” → the division is a statistical bias; many mixed-function sites exist on both sides
6. Corticospinal Tract — The Output Cable (5 min)
Core Message
The corticospinal tract is the main physical pathway from M1 to the spinal cord — but it’s not just M1.
Key Numbers (worth knowing for the exam)
- Origins: M1 (~40%), Premotor Cortex (~30%), Somatosensory Cortex (~20%)
- Contains ~1 million fibres in humans
- Only excitatory fibres (glutamatergic)
- Route: corona radiata → internal capsule → cerebral peduncles → pons → pyramids
- ~90% decussate at pyramidal decussation → contralateral limb control
- Forms lateral (crossed, 90%) and anterior (uncrossed, 10%) corticospinal tracts
Betz Cells — The Superstars
- Named for Volodymyr Betz (1834–1894); found only in Layer V of M1
- Largest neurons in the CNS — up to 100 μm in diameter
- Axons reach >1 m in humans (M1 to lumbar spinal cord)
- Directly contact α-motoneurons of brainstem and spinal cord
- Only ~5% of all cells projecting to spinal cord but most crucial for fast motor signals
- Conduction rate >70 m/sec — fastest brain-to-spinal cord pathway
Discussion Questions
-
“The corticospinal tract comes 40% from M1, 30% from premotor, 20% from S1. Why would somatosensory cortex send fibres down the corticospinal tract — what could a sensory area possibly be doing with direct spinal access?”
(Guide: S1 fibres modulate sensory gating in the spinal cord — adjusting how much sensory information reaches higher levels during movement. Efference copy / gating of self-generated sensory signals.) -
“Betz cells make up only ~5% of corticospinal fibres but are considered the most crucial. What does that tell you about how to think about the other 95%?”
(Guide: the other 95% likely modulate interneurons and spinal circuits rather than directly commanding motoneurons — more modulatory, context-setting role. Betz cells = direct executive command.)
7. Population Vector — Reading the Direction Code (12 min)
Core Message
M1 neurons vote for their preferred movement direction. No single neuron is decisive — the population determines the output.
Explanation Approach (Georgopoulos et al., 1986)
Setup: Monkey moves a manipulandum to 8 targets. Record from many M1 neurons.
Step 1 — Cosine tuning: Each neuron fires maximally for one direction (preferred direction, d_i), with firing decreasing as a cosine of the angular difference. This is coarse coding — comparable to orientation tuning in V1.
Step 2 — Finding d_i: The preferred direction is defined as:
This is a non-linear step — you take only the direction of maximal response.
Step 3 — Population vector:
The weighted sum of all neurons’ preferred direction vectors (weighted by their activity) approximates the actual movement direction. This is linear.
Key properties:
- PV predicts movement direction before movement begins (delay-phase activity)
- Dynamic: computed every 10–20 ms → “neural trajectory” tracks the planned path in real time
- Accuracy improves with more neurons; decreases steeply until ~50 neurons, then slowly — still room for improvement in BCI decoding
Discussion Questions
-
“A single M1 neuron fires for ‘right’ movements — but also somewhat for ‘upper-right’ and ‘lower-right.’ It’s a noisy, broad signal. Why is broad tuning actually BETTER for a population code than sharp, precise tuning?”
(Guide: if every neuron fired only for exactly one direction, nearby directions would have no representation — you’d need infinitely many neurons. Broad tuning means many neurons contribute to any movement direction, averaging out individual noise. This is the advantage of coarse coding.) -
“The population vector points in the planned movement direction already during the delay phase — before the monkey moves. What does that tell you about M1’s role in movement? Is M1 just ‘executing’ or is it also ‘planning’?”
(Guide: challenges the sharp execution-only view of M1. M1 is part of the planning process, not just the final output stage. But: “execution is dependent upon other areas” — the signal is set up but gated by BG/thalamus.) -
“The motor illusion task (Schwartz et al. 2007) showed that M1 population vectors track the physical hand movement while PMv tracks the perceived movement. Where in the motor hierarchy does the switch from sensory to motor space occur?”
(Guide: between PMv and M1. PMv = perceptual/planning representation of the action; M1 = the actual motor command. This dissociation is only visible through population vectors — single-neuron recording can’t isolate it as cleanly.) -
“If you’re building a BCI and you can implant 100 electrodes in M1, what does the confidence interval curve (which plateaus after ~50 neurons) tell you about where to invest your hardware effort?”
(Guide: more electrodes help but with diminishing returns past ~50. The real gains come from better algorithms and adapting the user to the device, not just more electrodes. Also: electrode placement quality matters more than raw count after a threshold.)
8. Repetition Suppression & Hierarchical Organisation (8 min)
Core Message
The motor system is not just M1. The action representation is distributed across regions that encode different aspects of movement — kinematics, goals, outcomes. Repetition suppression lets us carve this up non-invasively.
Principle
When the same neurons are activated twice in a row, the fMRI BOLD response is suppressed (neurons habituate). By varying which feature of an action is repeated vs. novel, you can infer what each region encodes.
Why necessary: fMRI can’t see individual neurons — repetition suppression acts as a virtual microscope.
Grafton & de Hamilton (2007) — Three Experiments
| Experiment | Feature A | Feature B | What’s dissociated |
|---|---|---|---|
| Exp 1 | Goal-Object | Trajectory | Goal coding vs. kinematics |
| Exp 2 | Goal-Object | Grip | Goal vs. grip type |
| Exp 3 | Outcome | Means | What’s achieved vs. how it’s done |
Findings:
- Kinematics (trajectory, grip) → posterior parietal cortex / visual association cortex
- Goal-Object → anterior intraparietal sulcus + left ventral premotor cortex
- Outcome → bilateral inferior parietal lobule + right ventral premotor cortex
Hierarchical conclusion: Motor system decomposes actions as: kinematics < goals < outcomes — each level processed in a different region.
Discussion Questions
-
“In repetition suppression, if a brain area shows suppression for BOTH goal repetition AND trajectory repetition — what does that mean about what that area encodes?”
(Guide: it encodes both features — it can’t be selective for just one. The suppression means the neurons in that region represent the repeated element, regardless of which one it is.) -
“Repetition suppression is used because fMRI can’t see individual neurons. Could you use it with direct electrode recordings in a monkey — and would it give you different information?”
(Guide: yes, but you’d see individual neurons suppressing to their preferred stimulus — you’d get the same logic at single-cell resolution. With electrodes you could check the timing of suppression, which you can’t with fMRI’s slow BOLD. Different information, same logic.) -
“The motor system organizes actions hierarchically as kinematics → goals → outcomes. What does this suggest about what the motor cortex is actually trying to do — control muscles, or control outcomes?”
(Guide: the hierarchy goes beyond muscle control. The system is solving an inverse problem: given a desired outcome, derive the appropriate goals, the movement kinematics, and finally the muscle commands. Motor cortex is part of an outcome-oriented system.)
9. Brain-Computer Interface — From 0.5 bit/s to BrainGate (8 min)
Core Message
BCIs are the clinical and technological translation of population vector coding from M1. The field went from non-invasive EEG spelling (1999) to implanted 100-electrode arrays (2006) and is still rapidly evolving.
Birbaumer et al. (1999, Nature) — How It All Started
- First non-invasive BCI: EEG (slow cortical potentials)
- Patient: locked-in syndrome — completely paralyzed, communicated only through eye movement
- Method: binary decisions — patient modulates cortical negativity vs. positivity to select A–M vs. N–Z, repeatedly halving the alphabet over 5–6 steps per letter
- Speed: ~25 seconds per letter → 0.5 bit/sec
- “Not much, but MUCH more than 0 bit/s” — the clinical significance was enormous
- Patient typed a letter to the experimenter — the first time communication was restored
BrainGate 2006 — Invasive Parallel Recording
- 100-electrode sensor (10×10 grid); 96 electrodes available for recording
- Electrode specs: 1 mm long, 400 μm spacing
- Implanted in pre-central knob (arm/hand “knob” area of M1 visible on MRI)
- First patient: tetraplegic, mechanically ventilated; controlled cursor in 16-target grid task using neural signals alone
- Bit rates higher than EEG; “massive parallel recording technology”
- “The promise is of huge improvements” — and indeed, current systems have made enormous progress since 2006
Discussion Questions
-
“Birbaumer’s system uses slow cortical potentials from EEG — not population vectors, not individual neurons. Yet it works. What does that tell you about how much neural precision you actually need for a BCI to be functional?”
(Guide: for binary decisions, you don’t need fine motor direction coding — just a coarse modulation signal. The algorithm can be simple if the task is simple. Full motor control requires the precision of M1 population vectors. There’s a trade-off between invasiveness and resolution.) -
“The BrainGate system implants 100 electrodes in M1. If electrodes degrade over months (glia scar formation), fewer neurons are recorded over time. How would that affect decoding — and how does the confidence interval curve from Georgopoulos 1988 help you think about this?”
(Guide: signal degrades, but the CI curve shows that up to ~50 neurons there’s a steep drop in precision — below 50 electrodes things get bad fast. Also: the decoder can be retrained as the neural population changes. This is a major real-world BCI engineering challenge.) -
“EEG-based BCI (non-invasive, 0.5 bit/s) vs. implanted array (invasive, much higher bit rate). You’re designing a BCI for someone with ALS who can still move their eyes. What approach would you choose and why?”
(Guide: ethical and practical. If a patient can still move eyes: eye-tracking BCI might be better than any neural BCI. EEG is safer but slower. Implanted array is faster but requires surgery, carries infection risk, degrades. For ALS specifically: disease progresses, eye movement may also be lost — argument for implant to future-proof. No clear right answer — the discussion is the point.)
10. Exam Question Round (12 min)
Select 3–4 from the 11 official exam questions based on what the group found hardest. Full list:
Q1. “Name 4 cortical areas of the motor system, give a simplified description of their function, and their relation to the respective Brodmann areas.”
Expected key points:
- M1 (BA4): executes movement, corticospinal output, Betz cells
- SMA (BA6 medial): self-initiated preparation, procedural memory; lesion → mutism
- Premotor (BA6 lateral): externally-guided movement, grasp patterns
- FEF (BA6+8): voluntary saccades, spatial attention
- Both SMA and PM are BA6 but distinct in location and function
Typical mistakes: Saying “SMA = premotor” without distinguishing them; wrong BA for FEF
Q2. “What can be said about the match of histological divisions and functional divisions in primary cortical areas and higher ones?”
Expected key points:
- Primary areas (M1, S1): histological (cytoarchitectonic) boundaries match functional boundaries well
- Higher areas (SMA, PMC, FEF): histological boundaries are rough guides only — functional divisions are finer and context-dependent
- M1/S1 boundary is “more of a bias than absolute” — mixed representations at the central sulcus
Q3. “What are the characteristics of the laminar structure of primary motor cortex?”
Expected key points:
- Agranular cortex — Layer 4 small or absent (no large thalamic input layer, unlike S1)
- Prominent Layer V: Betz cells (largest neurons in CNS, 100 μm, axons >1m, directly contact α-motoneurons, >70 m/sec, only ~5% of CST)
- Layer III pyramidal neurons: corticocortical connections
- GABAergic interneurons throughout all layers
Q4. “How does M1 project to and receive projections from other components of the motor system? Include non-cortical components.”
Expected key points:
- Receives from: Thalamus VL (from BG via GPi), SMA, Premotor, S1
- Projects to: Spinal cord (CST: 40% from M1, 30% PM, 20% S1), Brainstem motor nuclei, Reticular Formation
- CST: ~1M fibres, only excitatory, 90% cross at pyramidal decussation, forms lateral + anterior tracts
- Also projects to brainstem for head/neck motor control
Q6. “How does repetition suppression work? Describe one experiment. Why is it necessary?”
Expected key points:
- Principle: repeated activation of same neurons → BOLD suppression
- Necessary: fMRI can’t resolve individual neurons — RS acts as virtual super-resolution
- Grafton & de Hamilton (2007): 3 experiments crossing goal/trajectory/grip/outcome
- Findings: kinematics → parietal; goal-object → ant. IPS + left ventral premotor; outcome → inf. parietal lobule + right ventral premotor
Q9. “How could a movement be read out of M1 neurons? State and explain the population vector algorithm.”
Expected key points:
- Each M1 neuron has a preferred direction d_i = argmax_o(A_i(o)) — non-linear step
- Population vector P = Σ A_i(o_u) · d_i ≈ u — linear weighted sum
- Accuracy increases with neuron number; predictive even before movement onset (delay phase)
- Application: BCI decoding in real time
Q10. “Describe an experiment disentangling the role of premotor and motor cortex with a visual illusion.”
Expected key points:
- Schwartz et al. (2007): monkey draws 5 oval cycles in VR; gain ×1.8 applied cycles 3–4 → hand adapts to circle but perceived trajectory stays oval
- M1 population vector → tracks physical hand movement (circles)
- PMv population vector → tracks perceived movement (oval)
- Conclusion: switch from sensory to motor space occurs between PMv and M1
Q11. “When and how was BCI first invented and clinically applied?”
Expected key points:
- Birbaumer et al. (1999, Nature): EEG-based, slow cortical potentials; locked-in patient; binary alphabet halving; 0.5 bit/s; 25 sec/letter
- BrainGate (2006): 100 electrode implant in pre-central knob; tetraplegic patient controlled cursor; invasive; higher bit rate
- Principle: M1 population vectors decoded in real time to control external device
Closing (3 min)
Take-Home Message:
“The motor cortex is not a button. It’s a population vote. Thousands of neurons each vote for their preferred direction, and the movement that emerges is the vector sum of all those votes — computed every 20 milliseconds, anticipatory, plastic, and hierarchically organized from PMv (what you perceive) down to M1 (what you execute). When we record that vote, we can read the brain’s intended movement — that’s the promise of BCIs.”
Preview of next week (Lecture 5 — Mirror Neurons):
We’ve seen how M1 encodes what you do. But what happens in the motor system when you watch someone else move? Mirror neurons in premotor cortex (F5, the area corresponding to PMv) fire both when you act and when you observe an equivalent action.
Connecting question to leave them with:
“If your premotor cortex activates when you watch someone else move — does that mean understanding someone else’s action is fundamentally a motor process? And what would go wrong if that system were disrupted?”
Quick Reference — Key Numbers
| Fact | Value |
|---|---|
| Corticospinal fibres in humans | ~1 million |
| CST origin: M1 / Premotor / S1 | ~40% / 30% / 20% |
| CST decussation at medullary pyramids | ~90% |
| Betz cell diameter | up to 100 μm |
| Betz cell axon length | >1 m (M1 to lumbar cord) |
| Betz cell conduction velocity | >70 m/sec |
| Betz cells as % of CST | ~5% |
| Georgopoulos targets | 8 directions |
| Dynamic PV resolution | 10–20 ms |
| Birbaumer 1999 BCI speed | 0.5 bit/sec |
| BrainGate electrodes | 100 (10×10 grid); 96 available |
| BrainGate electrode spacing | 400 μm |